
by Richard Peachey
BEFORE 2002: When evolutionists discussed life on the early Earth, they could point with confidence to a variety of apparently well-documented evidence from several locations around the globe (Australia, Greenland, South Africa).
TODAY: Every piece of alleged evidence has now been thrown into doubt, and the field is in disarray. The current uncertainty is illustrated by the hesitant wording of one recent claim to have discovered ancient life on Earth: “This is the oldest sign of life that hasn’t been discredited, at least not yet [!]”1
[Note: A glossary of technical terms can be found at the end of this article.]
.
.
Australia: “Cellular” Microfossils
Our story begins in 1980 with the announcement by J. William Schopf of “5 microbial life forms in rock dated to 3500 million years old in the ‘North Pole’ region of northern Western Australia.”2 Schopf and his colleagues identified these as “unquestionable microfossils from the early Precambrian era.”3 Seven years later, Schopf and his graduate student Bonnie Packer reported:
“Cellularly preserved filamentous and colonial fossil microorganisms have been discovered in bedded carbonaceous cherts [flint-like sedimentary rock] from the Early Archean Apex Basalt and Towers Formation of northwestern Western Australia. . . . These fossils are among the oldest now known from the geologic record; their discovery substantiates previous reports of Early Archean microfossils in Warrawoona Group strata. . . . Together with other relevant data, they establish that microbial communities were extant, morphologically varied, and possibly physiologically advanced as early as 3.3 billion to 3.5 billion years ago.”4
In 1993 Schopf supplemented and reinforced his claims for the Australian microfossils:
“Eleven taxa (including eight heretofore undescribed species) of cellularly preserved filamentous microbes, among the oldest fossils known, have been discovered in a bedded chert unit of the Early Archean Apex Basalt of northwestern Western Australia. This prokaryotic assemblage establishes that trichomic cyanobacterium-like microorganisms were extant and morphologically diverse at least as early as ~3465 million years ago. . . . The Apex filaments meet all criteria required of bona fide Archean microfossils. . . . their evident cellular organization, and their morphological complexity and similarity to younger prokaryotes, both fossil and modern, firmly establish their biogenicity. . . . the morphological similarity of the Apex fossils to septate filamentous prokaryotes, both Proterozoic and modern, indicates that they are almost certainly prokaryotes and part of an evolutionary continuum that extends from the Early Archean to the present.”5
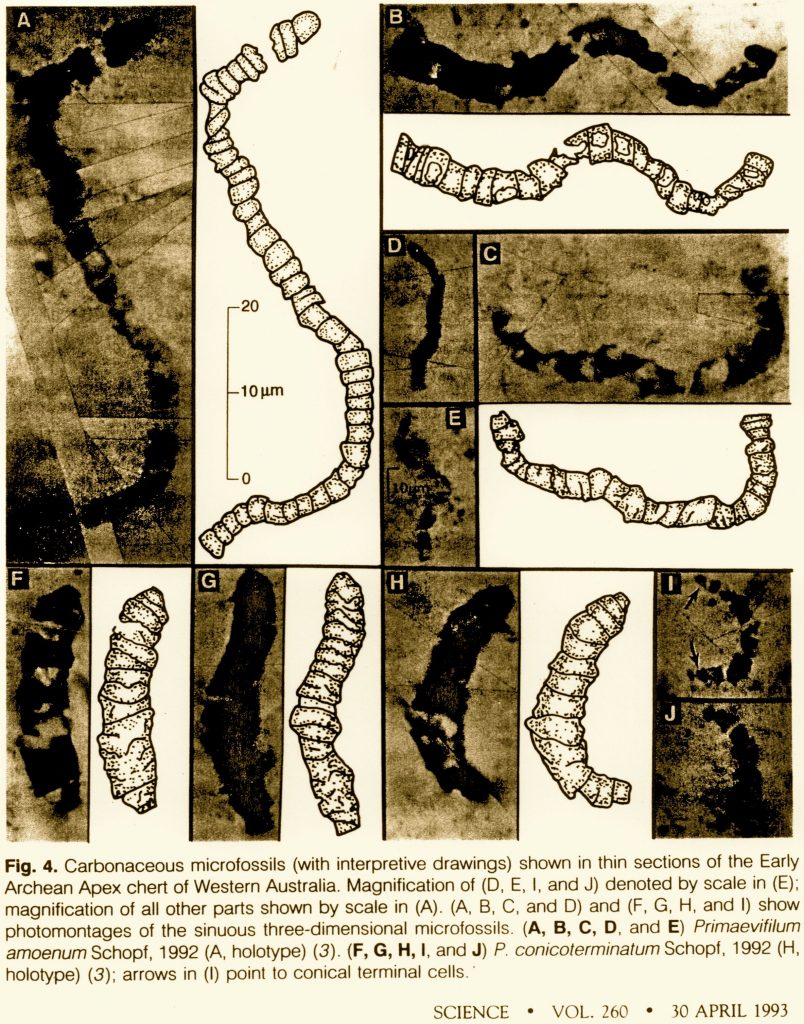
Alleged “microfossils” from J. William Schopf’s 1993 article
A few scientists were initially skeptical of such claims.6 “But over the years, Schopf’s description became textbook orthodoxy.”7 In a major technical review article, evolutionary “origin of life” researchers Stanley Miller and Antonio Lazcano commented as follows:
“The sediments in the Australian Warrawoona formation 3.5 x 109 years old contain very convincing cyanobacteria-like microfossils (Schopf, 1993). . . . There is compelling paleontological evidence that microbial communities were thriving on the primitive Earth 3.5 x 109 years ago (Schopf, 1993). . . .”8
Other evolutionists who bought into Schopf’s claim of “unquestionable microfossils” include Cambridge paleobiologist Simon Conway Morris, who described them as “convincing fossils in the form of minute cells,”9 and noted anti-creationist biology professor Massimo Pigliucci, who stated as fact that “the fossil record shows completely formed, ‘modern-looking’ bacterial cells a few hundred million years after the formation of Earth—about 3.5 billion years ago.”10
The Australian microfossil claims unraveled in 2002, thanks to the efforts of Oxford paleobiologist Martin Brasier. In an article critical of Schopf’s work, Brasier and his colleagues wrote:
“Eleven species of filamentous prokaryote, distinguished by shape and geometry, have been put forward [by Schopf] as meeting the criteria required of authentic Archaean microfossils, and contrast with other microfossils dismissed as either unreliable or unreproducible. . . . Here we report new research on the type and re-collected material, involving mapping, optical and electron microscopy, digital image analysis, micro-Raman spectroscopy and other geochemical techniques. We reinterpret the purported microfossil-like structure as secondary artefacts formed from amorphous granite within multiple generations of metalliferous hydrothermal vein chert and volcanic glass. . . . there is no support for primary biological morphology. . . .”11
Scientists in this field have by and large come to agree with Brasier; they have weighed Schopf’s work in the balance and found it wanting. And to add insult to injury, Schopf’s former graduate student Bonnie Packer has come forward to accuse him of deliberately concealing evidence contrary to his conclusions.12 The world-famous Australian discoveries remain under a cloud to this day.
Greenland: “Chemo-” Fossils
Photosynthetic organisms (such as algae and plants) preferentially incorporate carbon-12, the lighter of the two most common naturally-occurring isotopes of carbon. It has been theorized, therefore, that carbonaceous inclusions within minerals showing unusually high ratios of carbon-12 to carbon-13 would indicate the activity of once-living organisms even where no recognizable (cellular) fossils were found.13 In 1996, researchers who employed this approach at two Greenland sites announced enthusiastically:
“Here we report ion-microprobe measurements of the carbon-isotope composition of carbonaceous inclusions within grains of apatite (basic calcium phosphate) from the oldest known sediment sequences—a ~3,800-Myr-old banded iron formation from the Isua supracrustal belt, West Greenland, and a similar formation from the nearby Akilia island that is possibly older than 3,850 Myr. The carbon in the carbonaceous inclusions is isotopically light, indicative of biological activity; no known abiotic process can explain the data. . . . Together with the intergrowth of carbonaceous matter with apatite in BIF [banded iron formation] from Akilia island, we conclude that the isotopic results reported here give strong evidence for life on Earth by 3,850 Myr.”14
The first of the two Greenland sites to be challenged was Akilia island, in May 2002:
“On the island of Akilia, . . . southwest Greenland . . ., a sequence of lithologies that has been interpreted as mafic volcanic rocks intercalated with silicious sedimentary rocks chemically precipitated from seawater [banded iron formation, BIF] contains carbon isotopic signatures that have been interpreted as perhaps the oldest known life on Earth [>3850 million years ago (Ma)]. . . . Here we present new geologic, petrologic, and geochemical evidence that favors a metasomatized ultramafic igneous origin for rocks previously considered to be BIFs, indicating that it is highly improbable that the rocks hosted life at the time of their formation. . . . On the basis of our observations and data, we propose that the quartz-pyroxene rocks on Akilia originated as ultramafic intrusions or volcanic rocks, and therefore the carbon isotope signature of contained graphites is not indicative of past life. . . . Given these constraints, the best documented evidence for the earliest life on Earth is 13C-depleted graphite particles in deep-sea clastic sedimentary rocks from the Isua greenstone belt at 3700 to 3800 Ma. . . .”15
Within months the Isua site was also under fire — despite that hopeful comment at the end of the previous quote!
“Earlier studies on isotopic characteristics of graphite [carbon] occurring in rocks of the approximately 3.8-Gyr-old [i.e., 3.8-billion-year-old] Isua supracrustal belt (ISB) in southern West Greenland have suggested the presence of a vast microbial ecosystem in the early Archean. This interpretation, however, has to be approached with extreme care. Here we show that graphite occurs abundantly in secondary carbonate veins in the ISB that are formed at depth in the crust by injection of hot fluids reacting with older crustal rocks (metasomatism). . . . These metasomatic rocks, which clearly lack biological relevance, were earlier thought to be of sedimentary origin and their graphite association provided the basis for inferences about early life. The new observations thus call for a reassessment of previously presented evidence for ancient traces of lifein the highly metamorphosed Early Archean rock record. . . . It is now widely accepted that most, if not all, carbonate in Isua is metasomatic and not sedimentary in origin. . . . Our study leads to conclusions that require basic modification of some earlier inferences about traces of life in the ancient Isua rocks. A biogenic origin of graphite in carbonate-rich rocks in Isua was inferred from the assumption that these rocks had a sedimentary origin. However, recent field and laboratory investigations have shown that most if not all carbonate in Isua is metasomatic in origin.”16
Thus, what was once trumpeted as “strong evidence” has now been exposed as dubious “interpretation” or “inferences.” Nor has the Greenland situation improved since the criticisms published in 2002. Aivo Lepland, a coauthor of the article quoted above, has advised that after five years of work he and his colleagues have not been able to replicate the part of the original study related to Akilia island. And Gustaf Arrhenius, the senior author of the 1996 report (ref. 14), suspects now that there was “a mix-up of samples“(!)17
South Africa: Doubtable Dates
A 1991 survey of the “origin of life” field indicated that evidence for early life on Earth included “two sets of fossils from sites in Australia and South Africa whose age has been established through radioactive dating.” The South African material, said the report, “consists of lumpy, greenish-brown rocks that were once stromatolites, dense mounds of microbes that still grow in warm, shallow water in several locations around the world.”18 Five years later, researchers reported new findings related to a site in South Africa:
“We know life existed at least as far back as ~3.5 Ga [i.e., 3.5 billion years ago], as evidenced by microfossils occurring in cherts from various Archean greenstone belts (e.g., . . . Schopf, 1993). . . . we have chosen to test the hypothesis of life originating at sea-floor vents by focusing our attention largely on the organic signature of ancient vents contained within the Archean Barberton greenstone belt [South Africa]. . . . ironstone pods formed as constructional hydrothermal mounds directly on the sea floor and thus have a genesis similar to that of black smoker vents. . . . our results support the premise that sea-floor hydrothermal vents have always sustained vigorous organic activity and that diverse ecosystems, not unlike those within the oceans today, already existed 3.2 b.y. ago. Furthermore, the mass spectrometry patterns for organic compounds found within 3.2 Ga sea-floor hydrothermal vent samples are remarkably similar to those for modern vents . . . suggesting that ‘highly evolved’ microorganisms were already in existence ca. 3.2 Ga.”19
More recently, however, that conclusion has been brought into serious question:
“Over the past two decades, there has been a growing inference that hydrothermal processes played a major role in the chemical and biological evolution of early Earth. . . . One of the starting points for these arguments is the interpretation that iron oxide bodies, termed ‘ironstone pods,’ in the 3.5-3.2 Ga Barberton greenstone belt, South Africa, formed as deposits around submarine hydrothermal vents. . . . These bodies have provided what is arguably key evidence about life on Earth at 3230 Ma, including what are putatively the oldest-known complex organic compounds and evidence that Archean seafloor hydrothermal vents were populated by ‘”highly evolved” microorganisms’ (de Ronde and Ebbesen, 1996, p. 794). . . . We summarize evidence that the ironstone formed through young—Cenozoic and probably Quaternary—groundwater and/or low-temperature hydrothermal activity. . . . ironstone pods in the Barberton greenstone belt are not Archean in age and provide no information about early surface environments, ocean composition, or biological communities.”20
South Africa: One More Try!
In 2004 a team of “ancient life” researchers including Harald Furnes made it clear that they had not lost interest in South Africa:
“Pillow lava rims from the Mesoarchean Barberton Greenstone Belt in South Africa contain micrometer-scale mineralized tubes that provide evidence of submarine microbial activity during the early history of Earth. The tubes formed during microbial etching of glass along fractures, as seen in pillow lavas from recent oceanic crust. The margins of the tubes contain organic carbon, and many of the pillow rims exhibit isotopically light bulk-rock carbonate δ13C values [i.e., relatively low carbon-13 to carbon-12 ratios], supporting their biogenic origin. Overlapping metamorphic and magmatic dates from the pillow lavas suggest that microbial life colonized these subaqueous volcanic rocks soon after their eruption almost 3.5 billion years ago. . . . Our study indicates that microbes colonized basaltic glass of the early oceanic crust, much in the same way as they do modern volcanic glass. Well-preserved pillow lavas, which are a major component of Archean greenstone belts, may thus be one of the most promising places to search for vestiges of early life on Earth.”21
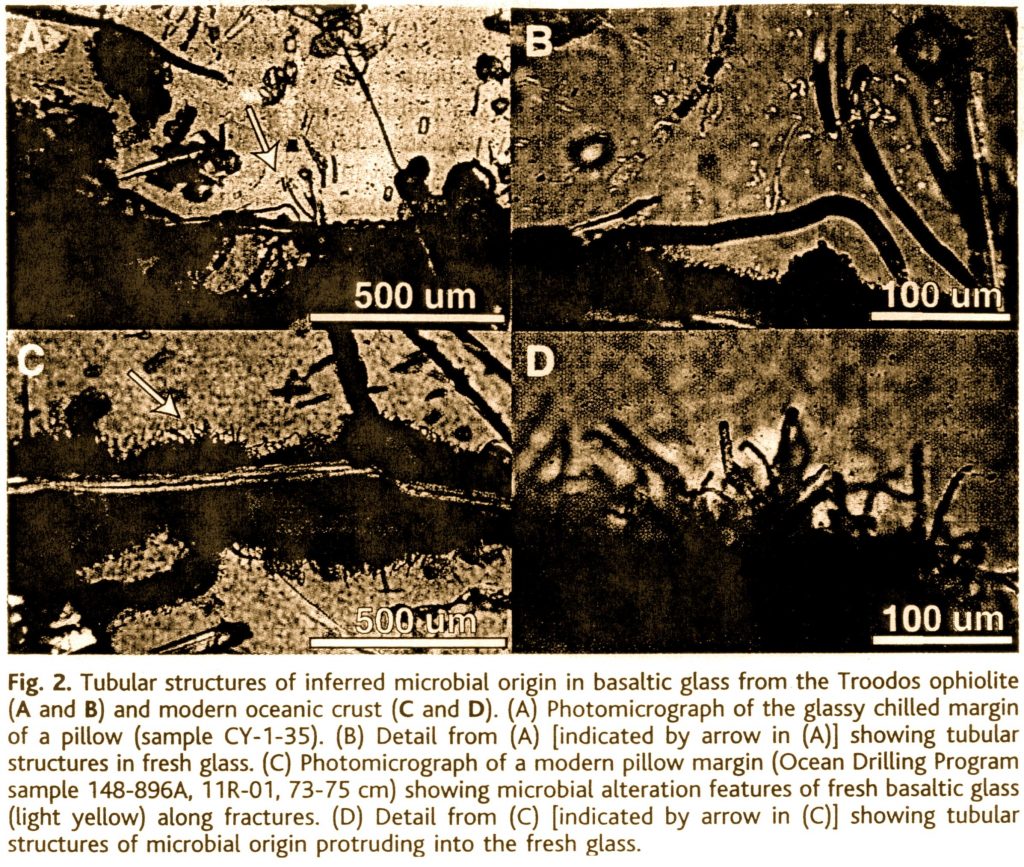
Alleged “microfossils” from Furnes et al.’s 2004 article
But this time the skeptics involved themselves early, wasting no time in questioning the claim:
“Unaware of Furnes’s latest work, [paleontologist Martin Brasier and his colleagues at the University of Oxford] were pursuing microtubules in Australian sea-floor lavas of the same age. ‘There’s no doubt theirs and ours are the same sort of thing,’ says Brasier, but ‘we have a completely different interpretation of what’s going on.’ In analogy to examples previously reported by others, Brasier believes that decomposition—possibly of organic matter—produced fluids that drove a mineral grain into the glass as chemical reactions at the grain ate into the glass, sort of like a corrosive-tipped pile driver. ‘We still do not know whether life is needed,’ says Brasier.”22
One of the researchers, University of Alberta geologist Karlis Muehlenbachs, reportedly stated: “This is the oldest sign of life that hasn’t been discredited, at least not yet.“23 That is perhaps the best that can be said for this latest claim!
Conclusion: The field’s current disarray as recently summarized in Nature
“Efforts to detect faint traces of life in Earth’s oldest rocks, some 4 billion years old, and in martian rocks that have fallen to Earth, have . . . produced results that are both ambiguous and disputed; claims of error in procedure and interpretation fly back and forth with regularity. . . . Few researchers now believe that ALH84001 [the ‘Mars rock’] is good evidence for the presence of past life on Mars. . . . Similar, and sometimes acrimonious, debates swirl around the evidence for the oldest life on Earth. Whether or not an outcrop of rock in Greenland contains evidence of life from 3.8 billion years ago is hotly contested. And the question of whether structures in 3.5-billion-year-old rocks in Australia are fossilized bacteria or artefacts produced in hot springs has been controversial for several years. . . . On Earth, living things preferentially incorporate the lighter form of carbon. . . . But, again, it is not clear how geology and chemistry influence isotope ratios, particularly in rocks that may be billions of years old. . . . Geological processes that transform carbonate minerals into organic matter seem to produce compounds that, like biological remains, are enriched with the lighter form of carbon, says geochemist Mark van Zuilen of the Petrographic and Geochemical Research Centre in Nancy, France. ‘I don’t think that carbon isotope ratios are that definitive an indicator of life,’ he says. ‘We probably don’t know all the ways in which isotope fractions can come about,’ adds geologist John Parnell of the University of Aberdeen, UK. ‘There may be things going on that we haven’t thought of yet.‘”24
Postscript
Several years after I assembled the above information, there has been yet another indication that supposed ancient microfossils are often nothing more than inorganic material: “Filamentous figments in the Apex Cherts,” published online by Nature, Feb. 20, 2011 <http://www.nature.com/news/2011/110220/full/news.2011.110.html>.
The news item begins, “Twenty years ago the palaeontological community gasped as geoscientists revealed evidence for the oldest bacterial fossils on the planet. Now, a report in Nature Geoscience shows that the filament structures that were so important in the fossil descriptions are not remnants of ancient life, but instead composed of inorganic material.”
The article quotes Oxford paleobiologist Martin Brasier as saying that people wanted to find life so badly that they ignored the obvious: “There is a willful blindness about these structures that sometimes has more to do with local politics than global truth.”
Glossary of Technical Terms
Archean Eon: “3.8 to 2.5 billion years ago.” (Early Archean: “3.3 to 3.5 bya.”)
Proterozoic Eon: “2500 to 544 million years ago.” [Dates given according to conventional evolutionary view.]
Igneous rocks are broadly categorized as felsic or mafic.
Felsic minerals (e.g., quartz, feldspars) are generally high in silica (SiO2), aluminum, potassium, and sodium. They have low melting points but are highly resistant to weathering. Rocks formed from felsic minerals generally are lighter in colour and are less dense than mafic mineral rocks.
Mafic minerals (e.g., mica, amphibole) are lower in silica, and high in magnesium and iron. They have high melting points but are less resistant to weathering. Rocks formed from mafic minerals are darker in colour and of greater density than felsic mineral rocks.
Ultramafic minerals (e.g., pyroxenes, olivine) are similar to mafic minerals but with very little or no silica.
Pyroxenes are a widespread group of dark green or grey rock-forming silicate minerals with the formula XYSi2O6 where X = Ca2+, Na+, Mg2+, Fe2+, Mn2+, or Li+, and Y = Mg2+, Fe2+, Mn2+, Fe3+, Al3+, Cr3+, or Ti4+.
Apatite, the principal phosphate mineral, is basic calcium phosphate, with the formula Ca5(PO4)3(F,Cl,OH) [mostly F, hence “fluorapatite”].
Pillow lava is lava (molten rock that has reached the Earth’s surface) ejected underwater to form pillow-shaped mounds of rock.
Greenstone refers to various kinds of compact igneous rock (“old lava”) that have taken on a characteristic green colour due to the development of chlorite, hornblende, or epidote.
A banded iron formation (BIF) consists of mafic volcanic rocks intercalated with silicious sedimentary rocks chemically precipitated from seawater.
Metasomatic rock is rock that has been changed in its bulk composition and mineral assemblage, usually by hot fluids penetrating it. The phenomenon is called metasomatism and the original rocks are said to have been metasomatized.
The term hydrothermal relates to the outpouring or ejection of hot water. A hydrothermal system is any natural system involving the heating and circulation of underground water and steam, powered by nearby hot or molten rocks. A hydrothermal vent is an outlet for heated water from a hydrothermal system, especially one at the bottom of an ocean. When the heated water is coloured with dark material, sulfides, and other minerals precipitated when the hot fluid comes into contact with cold seawater, the vent is known as a black smoker. Black smoker vents are thus associated with volcanogenic massive sulfide (VMS) deposits. They form at oceanic ridges and disgorge water about 400° C.
Ironstone pods are iron-oxide-rich mounds formed on the sea floor by hydrothermal activity.
The Gunflint Formation of Minnesota and Ontario is a succession of peculiarly banded silica- and iron-rich rocks: iron oxide (hematite) alternating with layers of red, yellow and grey chert.
References
1. Geologist Karlis Muehlenbachs, as quoted in Margaret Munro. 2004 (Apr 23). “Researchers: We’ve found earliest sign of earthly life.” The Province, p. A31. (Note: bold print within quotations given in this article represents emphasis added.)
2. As celebrated in: Norris D. McWhirter (ed.). 1985. Guinness Book of Records. 31st edition. Enfield, Middlesex, UK: Guinness Books. p. 53.
3. David I. Groves, John S. R. Dunlop, and Roger Buick. 1981 (Oct). “An Early Habitat of Life.” Scientific American 245(4):73.
4. J. William Schopf and Bonnie M. Packer. 1987 (Jul 3). “Early Archean (3.3-Billion to 3.5-Billion-Year-Old) Microfossils from Warrawoona Group, Australia.” Science 237:70, 72.
5. J. William Schopf. 1993 (Apr 30). “Microfossils of the Early Archean Apex Chert: New Evidence of the Antiquity of Life.” Science 260:640, 642f.
6. For example, Australian geologist Roger Buick. See John Horgan. 1991 (Feb). “In the Beginning . . .” Scientific American 264(2):120.
7. Rex Dalton. 2002 (Jun 20). “Squaring up over ancient life.” Nature 417:783.
8. Antonio Lazcano and Stanley L. Miller. 1996 (Jun 14). “The Origin and Early Evolution of Life: Prebiotic Chemistry, the Pre-RNA World, and Time.” Cell 85:793, 795. (The references within the quotation are to the article in ref. 5, above.)
9. Simon Conway Morris. 1998. The Crucible of Creation: The Burgess Shale and the Rise of Animals. Oxford, UK: Oxford University Press. pp. 22, 35.
10. Massimo Pigliucci. 1999 (Sep-Oct). “Where Do We Come From? A Humbling Look at the Biology of Life’s Origin.” Skeptical Inquirer 23(5):26 <http://web.utk.edu/~pteropus/pdfs/Pigliucci-1999-SI.pdf>. Yet another example of such (apparently unavoidable) credulity is the noted scientist Freeman Dyson of the Institute for Advanced Study, Princeton, who wrote: “I myself cannot pretend to decide whether a microscopic blob is a fossil cell or an ordinary grain of dust. I accept the verdict of the experts [N.B.!] who say that most of the blobs are in fact fossils.” Origins of Life, 2nd edition, 1999, p. 30 (Cambridge, UK: Cambridge University Press). Dyson pointed to South African “microfossils that resemble modern bacteria in shape and size” (p. 30) and urged that the Greenland chemo-fossil evidence “has to be taken seriously” (p. 32). Interestingly, just a page later (p. 33), Dyson offered this reality check concerning “experts”: “The experts [N.B.] believed that at the epoch of life’s origin the earth’s atmosphere was reducing and contained the hydrogen-rich species ammonia, methane, and molecular hydrogen that Miller used in his experiment. Miller’s experiment was supposed to be a true simulation of prebiotic chemistry on the primitive earth. But now nobody believes this any more.“
11. Martin D. Brasier, Owen R. Green, Andrew P. Jephcoat, Annette K. Kleppe, Martin J. Van Kranendonk, John F. Lindsay, Andrew Steele, and Nathalie V. Grassineau. 2002 (Mar 7). “Questioning the evidence for Earth’s oldest fossils.” Nature 416:76f. Brasier subsequently repeated and broadened his skepticism over putative fossils of early life: Martin Brasier, Nicola McLoughlin, Owen Green, and David Wacey. 2006. “A fresh look at the fossil evidence for early Archaean cellular life.” Philosophical Transactions of the Royal Society B 361:887–902 (available online at <http://physwww.mcmaster.ca/~higgsp/3D03/BrasierArchaeanFossils.pdf>). In 2011, Wacey, Brasier, and three coauthors claimed new and better evidence of cellular microfossils: David Wacey, Matt R. Kilburn, Martin Saunders, John Cliff and Martin D. Brasier. 2011 (Aug 21). “Microfossils of sulphur-metabolizing cells in 3.4-billion-year-old rocks of Western Australia.” Nature Geoscience 4:698-702. They claim: “For the first time in Archaean rocks [N.B.], we find a direct association between cellular morphology and metabolic by-products (micrometre-sized pyrite), and between cellular morphology and potential electron donors (detrital pyrite) under dysaerobic conditions. Taken together, these joint occurrences provide strong evidence of a multicomponent sulphur-based bacterial ecosystem preserved within this new ~3,400 Myr-old microfossil assemblage.” At the same time, they repeatedly acknowledge the problematic nature of such claims: “Determining the biogenicity of putative Archaean microfossils is notoriously difficult. No single line of evidence provides adequate proof of biological processing, instead, multiple lines of mutually supporting morphological and geochemical evidence are required. . . . Biology-like morphology and behaviour can seldom completely exclude an origin from abiotic artefacts, because abiotic carbonaceous coatings of grain margins may mimic cellular morphology. . . . Abiotic carbonaceous material, formed by Fischer-Tropsch-type synthesis or metamorphic reduction of siderite, could feasibly replicate one or more of the geochemical features described above. . . . such δ34S fractionations can occur during non-biological reactions, for example during the disproportionation of SO2 to sulphate and sulphide in oxidizing magmatic fluids. . . .” Wacey and colleagues think their evidence is strong enough to overcome this range of potential difficulties. Time will tell.
12. Rex Dalton, ref. 7, pp. 782-784. Doubts about Schopf’s work have remained strong: Robert Riding. 2008 (Feb 1). “Drifters Through Time.” Science 319:571.
13. Manfred Schidlowski. 1988 (May 26). “A 3,800-million-year isotopic record of life from carbon in sedimentary rocks.” Nature 333:313-318.
14. S. J. Mojzsis, G. Arrhenius, K. D. McKeegan, T. M. Harrison, A. P. Nutman, and C. R. L. Friend. 1996 (Nov 7). “Evidence for life on Earth before 3,800 million years ago.” Nature 384:55f., 59. Based on such “strong evidence,” Francis Collins, director of the Human Genome Project and theistic evolutionist, stated as fact that “3.85 billion years ago, life was teeming.” Francis S. Collins. 2003. “Faith and the Human Genome.” Perspectives on Science and Christian Faith 55(3):152.
15. Christopher M. Fedo and Martin J. Whitehouse. 2002 (May 24). “Metasomatic Origin of Quartz-Pyroxene Rock, Akilia, Greenland, and Implications for Earth’s Earliest Life.” Science 296:1448-1451. (Words in square brackets are the authors’ own.) See also the related story (in the same issue): Richard A. Kerr. 2002 (May 24). “Reversals Reveal Pitfalls in Spotting Ancient and E.T. Life.” Science 296:1384f. Kerr provides statements from other scientists in the field that are supportive of Fedo and Whitehouse’s conclusions. A team including Schopf subsequently attempted to vindicate “chemical fossils of early life” found in Akilia. Unlike Schopf’s earlier excessively confident approach, however, the researchers advise: “We do not claim that the data presented here constitute unequivocal evidence of early life.” Kevin D. McKeegan, Anatoliy B. Kudryavtsev, and J. William Schopf. 2007 (Jul). Geology 35(7):591-594.
16. Mark A. van Zuilen, Aivo Lepland & Gustaf Arrhenius. 2002 (Aug 8). “Reassessing the evidence for the earliest traces of life.” Nature 418:627, 629. More recently, these scientists have again concluded that “the Akilia apatite has no bearing on claims pertaining to a past record of life on Earth.” Aivo Lepland, Mark A. van Zuilen, Gustaf Arrhenius, Martin J. Whitehouse, and Christopher M. Fedo. 2005. “Questioning the evidence for Earth’s earliest life—AkiIia revisited.” Geology 33(1):77.
17. Rex Dalton. 2004 (Jun 17). “Fresh study questions oldest traces of life in Akilia rock.” Nature 429:688. In a more recent statement, an Oxford geologist lamented, “To my regret, the ancient Greenland rocks have not yet produced any compelling evidence for the existence of life by 3.8 billion years ago.” Stephen Moorbath. 2005 (Mar 10). “Dating earliest life.” Nature 434:155. Moorbath concludes his article: “For the time being, the many claims for life in the first 2.0-2.5 billion years of Earth’s history are once again being vigorously debated: true consensus for life’s existence seems to be reached only with the bacterial fossils of the 1.9-billion-year-old Gunflint Formation of Ontario.” Since claims for the earliest life had been connected with dates around 3.8 billion years ago, this Oxford geologist’s statement means that half of the fossil record, chronologically speaking, is currently up for dispute! Regarding fossils dated prior to 1.9 billion years ago, Harvard paleontologist Andrew Knoll can only offer: “A single fossil in 2.7-billion-year-old cherts from Australia looks tantalizingly but not unambiguously cyanobacterial, and 2.5-billion-year-old cherts from South Africa contain indifferently preserved remains that could be blue-greens, as well. Fortunately, better evidence of cyanobacterial antiquity comes from an unexpected source—biomarker molecules [i.e., not cells!] in 2.7-billion-year-old shales found just south of North Pole in northwestern Australia.” Andrew Knoll. 2005. Life on a Young Planet: The First Three Billion Years of Evolution on Earth. Princeton, NJ: Princeton University Press. p. 93.
18. John Horgan, ref. 6. Linking this South African material with Schopf’s claims, Harvard paleontologist Stephen Jay Gould stated positively and dogmatically: “The oldest rocks sufficiently unaltered to retain cellular fossils—African and Australian sediments dated to 3.5 billion years old—do preserve prokaryotic cells (bacteria and cyanophytes) and stromatolites (mats of sediment trapped and bound by these cells in shallow marine waters).” Stephen Jay Gould. 1994 (Oct). “The Evolution of Life on the Earth.” Scientific American 271(4):87.
19. Cornel E. J. de Ronde and Thomas W. Ebbesen. 1996 (Sep). “3.2 b.y. of organic compound formation near sea-floor hot springs.” Geology 24(9):791, 794. (The reference within the quotation is to the article in ref. 5, above.)
20. Donald R. Lowe and Gary R. Byerly. 2003 (Oct). “Ironstone pods in the Archean Barberton greenstone belt, South Africa: Earth’s oldest seafloor hydrothermal vents reinterpreted as Quaternary subaerial springs.” Geology 31(10):909, 912. (The reference within the quotation is to the article in ref. 19, above.) A comment on this article from de Ronde et al., with a reply from Lowe and Byerly, can be found at <http://geology.gsapubs.org/content/32/1/e68.full.pdf+html>. Note that Lowe and Byerly’s suggested age reduction is from 3230 million years ago to the Quaternary (which began 1.64 million years ago according to Peter W. Price. 1995. Biological Evolution. Fort Worth, TX: Saunders College Publishing. p. 122). This amounts to a revised age which is 1.64/3230 or about 1/20 of 1% of the originally proposed age. One might be caused to wonder about the accuracy of all the other dates found in the “scientific” literature!
21. Harald Furnes, Neil R. Banerjee, Karlis Muehlenbachs, Hubert Staudigel, and Maarten de Wit. 2004 (Apr 23). “Early Life Recorded in Archean Pillow Lavas.” Science 304:578, 580f.
22. Richard A. Kerr. 2004 (Apr 23). “New Biomarker Proposed for Earliest Life on Earth.” Science 304:503.
23. Margaret Munro, ref. 1.
24. John Whitfield. 2004 (Jul 15). “It’s life . . . isn’t it?” Nature 430:288f. By the way, a more recent discussion points out that there aren’t any promising locations to hunt for “early life” other than the ones we have treated in this article: “When searching for evidence of past life, sedimentary environments are considered the most suitable because they are often formed in association with water, a fundamental requirement for life. There are only three known locations that host exposures of ancient sediments: Isua and Akilia in southwest Greenland, which are [allegedly] 3.8 to 3.7 billion years old (Ga), the Pilbara in northwestern Australia (3.5 to 3.3 Ga), and Barberton in eastern South Africa (3.5 to 3.3 Ga).” Frances Westall. 2005 (Apr 15). “Life on the Early Earth: A Sedimentary View.” Science 308:366f. Geoscientists have continued to be cautious about the significance about carbon isotope ratios: Sid Perkins. 2008 (Aug 2). “Tiny diamonds may set earlier date for first life.” Science News 174(3):13.